

My research focuses on sexually heteromorphic traits; everything from genetics to hormones, evolutionary patterns, mating systems, and behaviors.
Cross-Sexual Transfer and The Ancestral Modulation Hypothesis
This is the overarching concept that drives research in my lab. Cross-sexual transfer is a concept proposed by West-Eberhard in 2003 that describes an evolutionary situation where trait(s) expressed in one sex become expressed in the other. This can result in secondary monomorphisms or new heteromorphic patterns and can occur across species, populations, individuals, or even lifetimes. Cross-sexual transfer may help account for sexual mosaicism, the concept that sexes are often heteromorphic for multiple phenotypes, but that within any given individual, each of these phenotypic traits can lie at different points of their respective population distributions.

A missing part of the story of cross-sexual transfer is the mechanistic patterns that govern these traits, which is where the ancestral modulation hypothesis comes in. I developed this concept to explain what mechanisms that regulate sexual heteromorphism might emerge as heteromorphism develops (I call Type I traits) and what mechanisms might govern cross-sexual transfers (Type II traits). While there are many ways of generating sex-bias, I focus on vertebrates and hormonal signaling systems. In short, a Type I Trait emerges through the gain of genomic elements that are responsive to already sex-biased hormonal signaling which therefore cause different expression patterns across sexes; whereas a Type II Trait does not change these genomic elements and instead alters the sex-bias signal to access these elements which allows for environmental responses and cross-sexual transfer to work as a plastic response.
Papers I have written on these subjects can be found Here (Cross-sexual transfer) and Here (Ancestral Modulation Hypothesis)
I focus my work on fishes because they are amazingly diverse in sexually heteromorphic traits such as ornamentation, broodcare, and behavior. While my questions focus on Cross-Sexual Transfer and Ancestral Modulation Hypothesis, I will follow new findings in whatever direction they lead. I am also open to working in other systems to test the applicability of Cross-Sexual Transfer and Ancestral Modulation Hypothesis.
Stay tuned for upcoming cricket work!
I am stepping away from both fishes and parts of the Ancestral Modulation Hypothesis and Cross-sexual transfer for the time being to work as a post-doc with Mingzi Xu. I am both going back and forward. I’m am going back into something I’ve always enjoyed: mate choice and variation; while also going forward with a brand new (to me) system: field crickets. No only am I combining these two pieces, but I am bringing them together with my view towards signaling variation by investigating variation in female choice, hormonal levels, and signaling pathway gene expression. I hope to share some more details as data is more fully collected.
Current Projects (* = Student Led)
Julidochromis and Changes in Mating Pair Dynamics
The cichlid genus, Julidochromis, has biparental care with a male/female pair cooperating to spawn and care for the eggs and fry in the nest. The larger animal in the pair mostly swims around the perimeter of the nest with the smaller animal staying in the nest and tending the fry more directly. In one species, pairs are usually male-larger (J. transcriptus) and in another species pairs are usually female-larger (J. marlieri), but in both species pairs can be manipulated to form opposite pairs where individuals take on the role based on size, not sex. This could be a case of cross-sexual transfer that occurs both within and across species as the behaviors could have evolved from a heteromorphic ancestor that likely engaged in male-biased territory defense. As a result, these “Julies”, as they are affectionately called, are a great system to study.
Experimental work has completed since I have departed Reed College; however, data, genetics, and genomic work is ongoing with results still being published
Julidochromis marlieri
Hormonal Changes with Shifting Roles*
Since Julies can change their roles in a pair during their lifetime, we are partnering individuals with larger and smaller mates and observing the behavior to confirm the presence and degree of changes in pair behaviors individuals engage in. Following the establishment and behavioral documentation of pairing behavior we sample gonadal steroid hormones (testosterone, 11-ketotestosterone, estradiol, and progesterone) so we can see how changes in partner size affects circulating hormones and how those correlate with documented behaviors. We are doing this in both species (transcriptus and marlieri) to see if the preferred mating dynamic has any affect on these parameters. The end result is a reaction norm of our responses (hormones and behaviors) with the individuals position in the pair as either the dominant territory guardian or submissive nest tender.
—Current results suggest J. transcriptus has an influx of 11KT when taking on the Guardian role in the nest, while no hormonal changes are present in J. marlieri. See paper in CV
Julidochromis transcriptus
Genetics and Shifting Roles
The hormonal experiment described above is one half of the overall work on these fishes. The other part is to extract the brains of these fishes at the end of the experiment and process them for transcriptomic analysis. Previous work in the Renn lab has demonstrated genetic expression overlap based on role in the pair between the two species (e.g. female marlieri and male transcriptus have similar expression patterns), but the plasticity within species has not been investigated. We will not be doing whole brain transcriptome, instead we will be focusing on and sampling regions of the brain associated aggression and broodcare in fishes, allowing us to have a clearer picture of the genes that correlate with the behavioral patterns seen in the pair. Further, by combining genetic and hormonal results with behavioral results we will have a more integrated picture of how these behaviors are regulated and how social environment and sex interact to generate these patterns.
Past Projects
Exogenous Hormones Influence on Behavior*
With some evidence pointing towards a correlation between circulating 11KT levels and aggressive behaviors, we are testing if the hormone is the causitive agent of change. We are piloting research by inserting an 11KT capsule into the body cavity of Julidochromis sp. and observing the changes in behavior. We will assay behavioral changes both within a large social setting and within a pair dynamic. Our aim is to have the 11KT-treated fish take on the behavioral role of a large fish even when it is equal or slightly smaller in size than it’s partner.
—We did see some behavioral changes following implantation, but we could not confirm any hormonal changes suggesting our technique needs more refinement and the surgery introduces significant confounders to behavior
Hybridization Between Congeners*
Looking at J. transcriptus and J. marlieri, there are few features to distinguish the species. We are interested if the animals can readily cue into these differences and respond to them. Ideally, an individual might hybridize with no other available options, which could pave the way for some exciting QTL studies. This is a long shot as these species likely evolved in sympatry and therefore have some strong pre-copulatory barriers in place. Indeed, other research has shown these fish can recognize and remember individuals by facial patterns.
—Our attempts, did indeed lead to a failure to hybridize and an exclusion of non-specifics. Behavioral data suggests J. transcriptus may be more resistant to pairing than J. marlieri though some possible courtship was observed between J. transcriptus males and J. marlieri females.
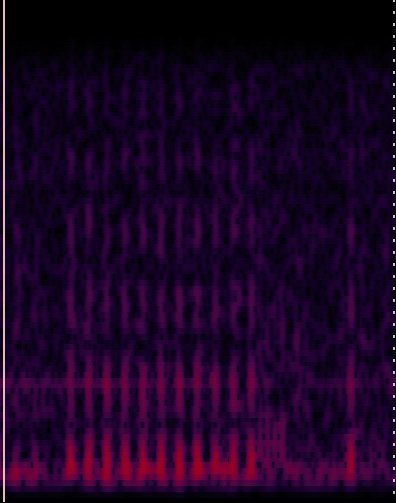
Spectrogram of J. transcriptus

Wave form of J. transcriptus
Singing Julies??*
Until recently, the soundscape of fishes was under-appreciated but there has been an increase in awareness that fishes communicate beyond sight and smell and more researchers are “mic-ing up” their fishes. We are no exception, our Julies engage in a range of behaviors within their pairs, but one that fascinates us is a “shimmy” they perform. This shimmy is similar to those seen in Astototilapia burtoni when males court females (and in other contexts). It turns out males sometimes generate a sound when engaging in this behavior so we want to know if our Julies make sounds as well and how those may differ across species, sexes, and various contexts. We have replicated the results in A. burtoni and are just now starting in our Julies!
Addendum: They do!!
Sphaerichthys osphromenoides
Rapid Chromosome Changes in a Gourami Subfamily*
In a group of gourami in the Luciocephalinae subfamily it is known that one species, Sphaerichthys osphromenoides, has an incredibly low chromosome count (2n = 16). We have been investigating other members of the genus and the next out to determine where the in the evolutionary history chromosome count drastically decreased. It turns out the karyotypes within this group vary widely. We have observed chromosome counts from 2n = 14 to 2n = 28 with sister species having very different numbers. We published these results with the hope to one-day do full genomic sequencing for synteny analysis.

Mouthbrooding A. burtoni
Female Competition During Mouthbrooding in A. burtoni*
Astotatilapia burtoni engage in female mouthbrooding of eggs and early fry. This maternal care is costly as the females do not eat during egg incubation and must guard their fry from being consumed by other fishes, including conspecifics. Given all these pressures, females must decide on continued investment in her offspring or cutting her loses and recouping energy (i.e. cannibalizing her brood) for future reproductive efforts. We are seeing how female-female social dynamics may affect this choice by putting pairs of females with similarly aged broods within sight of each other and occasionally allowing the females to physically interact. We are interested in how social standing can affect female decisions on parental effort.

Pair Dynamics and Behaviors in Julies in Response to an Intruder*
We have finished a project started previously in the Renn Lab on how pairs of J. marlieri respond to a conspecific intruder. We manipulated which sex was larger in the pair and the sex of the intruder. Results are in the process of being published, but in general there is a plastic response to and by intruders that is context-dependent based on the sex of the intruder and sex of the larger individual, though large individuals always show more interactions with intruders than smaller individuals.
Syngnathus scovelli females from Florida Keys
Role of Hormone Response Elements in Sex-biased Traits and Sexual Selection
This was the thrust of my doctoral work. Working in pipefish I confirmed the importance of female-specific vertical iridescent bands in male mate-choice. I expanded on the knowledge that these bands are estrogen-mediated, by showing that genes putatively involved in band formation had an excess of genomic regions that could be bound by estrogen receptors when compared to other gene sets. Further work demonstrated that human chromosomes with more estrogen-responsive genes have greater estrogen binding capabilities than chromosomes with less estrogen-responsive genes. I also did a comparative approach across primates showing that as some features associated with sexual selection such as canine size heteromorphism influence the genome-wide presence of androgen response elements. These findings got me started on my Ancestral Modulation Hypothesis as I suspect these response elements emerge as heteromorphic traits appear and are preserved during cross-sexual transfer due to the plastic nature of cross-sexual transfer. I am still in contact with collaborators to resume work on pipefishes and have plans to pursue future work into the variation in estrogen response elements within the ornamented species and across ornamented and unornamented species.